Résumé
Comprendre et anticiper la présence du cerf par la lecture du paysage
Comment anticiper la présence d'un animal sauvage dans un territoire inconnu ? Cette question constitue l'un des principaux défis de l'observation naturaliste et de la photographie animalière. À partir d'observations menées dans plusieurs régions de montagne suisses et confrontées aux résultats récents de l'écologie spatiale, de l'écologie du paysage et de la télémétrie GPS, cet article explore la manière dont l'interprétation du paysage peut aider à comprendre et à anticiper la présence du cerf élaphe (Cervus elaphus).
Les observations suggèrent que l'utilisation de l'espace dépend moins de parcelles d'habitat isolées que de leur intégration dans un système territorial plus large combinant connectivité, accessibilité, disponibilité des ressources et sécurité. Plusieurs motifs récurrents apparaissent dans des paysages contrastés : corridors de déplacement, interfaces entre milieux ouverts et couverts, gradients altitudinaux et zones refuges structurent la distribution du cerf.
Sur cette base, l'article propose une typologie simplifiée des territoires à cerfs, un cadre saisonnier décrivant les changements d'utilisation de l'espace au cours de l'année, ainsi qu'une discussion sur les différences de comportement spatial entre mâles et femelles. Il présente enfin une méthode pratique de lecture prédictive du paysage destinée à améliorer la prospection naturaliste.
Mots-clés : cerf élaphe, Cervus elaphus, écologie spatiale, écologie du paysage, télémétrie GPS, connectivité des habitats, corridors écologiques, déplacements de la faune, analyse prédictive du paysage, observation naturaliste, Suisse.
Abstract
Understanding and anticipating red deer presence through landscape interpretation
How can the presence of a wild animal be anticipated in an unfamiliar landscape? This question represents one of the main challenges of wildlife observation and nature photography. Drawing on observations conducted in several Swiss mountain regions and comparing them with recent findings from spatial ecology, landscape ecology and GPS telemetry studies, this article explores how landscape interpretation can help understand and anticipate the presence of red deer (Cervus elaphus).
The observations suggest that space use depends less on isolated habitat patches than on their integration within a broader territorial system combining connectivity, accessibility, resource availability and safety. Several recurring patterns emerge across contrasting landscapes, highlighting the importance of movement corridors, habitat interfaces, altitudinal gradients and refuge areas in structuring deer distribution.
Based on these observations, a simplified typology of red deer territories is proposed, together with a seasonal framework describing how landscape use changes throughout the year. Differences in spatial behaviour between males and females are also discussed. Finally, the article outlines a practical approach to predictive landscape reading aimed at improving wildlife prospecting and helping observers identify the sectors most likely to be used by deer. Beyond the specific case of red deer, this contribution illustrates how a geographical approach to landscape analysis can provide valuable insights into the spatial organisation of large mammals and support more effective field observation.
Keywords: red deer, Cervus elaphus, spatial ecology, landscape ecology, GPS telemetry, habitat connectivity, ecological corridors, wildlife movements, predictive landscape analysis, wildlife observation, Switzerland.
Zusammenfassung
Die Anwesenheit des Rothirsches durch Landschaftsinterpretation verstehen und vorausahnen
Wie lässt sich die Anwesenheit eines Wildtiers in einer unbekannten Landschaft abschätzen? Diese Frage gehört zu den zentralen Herausforderungen der Naturbeobachtung und der Wildtierfotografie. Auf der Grundlage von Beobachtungen in mehreren Schweizer Gebirgsräumen und im Abgleich mit neueren Erkenntnissen der Raumökologie, Landschaftsökologie und GPS-Telemetrie untersucht dieser Beitrag, wie Landschaftsinterpretation helfen kann, die Präsenz des Rothirsches (Cervus elaphus) zu verstehen und vorauszudenken.
Die Beobachtungen deuten darauf hin, dass die Raumnutzung weniger von isolierten Habitatflächen abhängt als von ihrer Einbindung in ein größeres territoriales System aus Vernetzung, Zugänglichkeit, Ressourcenverfügbarkeit und Sicherheit. Über sehr unterschiedliche Landschaften hinweg treten wiederkehrende Strukturen auf: Bewegungskorridore, Habitatübergänge, Höhengradienten und Rückzugsräume prägen die Verteilung der Hirsche.
Auf dieser Grundlage wird eine vereinfachte Typologie von Rothirschgebieten vorgeschlagen, ergänzt durch ein jahreszeitliches Modell der Raumnutzung und eine Diskussion der unterschiedlichen räumlichen Strategien von Männchen und Weibchen. Abschließend wird ein praktischer Ansatz für eine vorausschauende Landschaftslesung vorgestellt.
Schlüsselwörter: Rothirsch, Cervus elaphus, Raumökologie, Landschaftsökologie, GPS-Telemetrie, Habitatvernetzung, ökologische Korridore, Wildtierbewegungen, Schweiz.
Riassunto
Comprendere e anticipare la presenza del cervo attraverso la lettura del paesaggio
Come si può anticipare la presenza di un animale selvatico in un territorio sconosciuto? Questa domanda rappresenta una delle principali sfide dell'osservazione naturalistica e della fotografia faunistica. A partire da osservazioni condotte in diverse regioni montane svizzere e confrontate con risultati recenti dell'ecologia spaziale, dell'ecologia del paesaggio e della telemetria GPS, questo articolo esplora come l'interpretazione del paesaggio possa aiutare a comprendere e anticipare la presenza del cervo nobile (Cervus elaphus).
Le osservazioni suggeriscono che l'uso dello spazio dipende meno da parcelle di habitat isolate che dal loro inserimento in un sistema territoriale più ampio, che combina connettività, accessibilità, disponibilità di risorse e sicurezza. In paesaggi molto diversi emergono schemi ricorrenti: corridoi di movimento, interfacce ecologiche, gradienti altitudinali e aree rifugio strutturano la distribuzione dei cervi.
Sulla base di queste osservazioni viene proposta una tipologia semplificata dei territori del cervo, insieme a un quadro stagionale dell'uso dello spazio e a una discussione sulle differenze di comportamento spaziale tra maschi e femmine. L'articolo presenta infine un metodo pratico di lettura predittiva del paesaggio.
Parole chiave: cervo nobile, Cervus elaphus, ecologia spaziale, ecologia del paesaggio, telemetria GPS, connettività degli habitat, corridoi ecologici, movimenti della fauna, Svizzera.
Introduction
De l'observation directe à la lecture géographique du territoire
Lorsqu'un naturaliste découvre un territoire inconnu, une question revient rapidement : où chercher les animaux ? Cette interrogation est particulièrement importante en photographie animalière. Le temps disponible sur le terrain est souvent limité, tandis que les habitats potentiellement favorables peuvent couvrir plusieurs dizaines de kilomètres carrés. Dans un tel contexte, l'observation directe ne suffit pas toujours. Il devient nécessaire de développer une lecture du paysage permettant d'identifier les secteurs les plus prometteurs avant même la première observation.
Cette démarche se situe à l'intersection de la géographie et de l'écologie. Elle consiste à considérer un territoire non comme une simple juxtaposition d'habitats, mais comme un système spatial structuré par des gradients altitudinaux, des interfaces écologiques, des corridors de déplacement, des zones refuges et des connexions avec les paysages voisins. Les animaux deviennent alors non seulement des objets d'observation, mais aussi des indicateurs du fonctionnement territorial des paysages qu'ils occupent.
Les réflexions présentées dans cet article sont issues de plusieurs années d'observation et de photographie animalière réalisées dans différentes régions de montagne suisses. Avec le temps, certaines régularités sont apparues. Pourquoi certains pâturages semblent-ils régulièrement fréquentés alors que d'autres restent vides ? Pourquoi certaines vallées fonctionnent-elles comme des corridors de déplacement tandis que d'autres concentrent les animaux pendant de longues périodes ? Pourquoi des paysages géographiquement très différents contiennent-ils néanmoins des structures spatiales qui remplissent des fonctions comparables ?
Ces questions ont progressivement conduit à un changement de perspective. L'objectif n'était plus seulement de localiser les animaux, mais de comprendre les processus spatiaux qui organisent leur présence. Les cols, les lignes de crête, les interfaces forêt-milieu ouvert, les zones peu dérangées et les gradients altitudinaux sont peu à peu devenus aussi importants que les animaux eux-mêmes.
Le cerf élaphe (Cervus elaphus) constitue un modèle particulièrement intéressant pour explorer ces questions. L'espèce est aujourd'hui largement distribuée dans les massifs suisses, ce qui permet d'observer ses populations dans des contextes paysagers très variés. Elle reste en même temps suffisamment discrète et mobile pour rendre l'observation exigeante. Plus imprévisible que de nombreux autres grands herbivores de montagne, le cerf encourage l'observateur à dépasser la simple recherche visuelle et à considérer le fonctionnement du territoire dans son ensemble.
Les observations présentées ici ont été réalisées avant toute recherche bibliographique spécifique. Ce n'est qu'ensuite qu'elles ont été comparées aux résultats récents de l'écologie spatiale, de la télémétrie GPS et de l'écologie du paysage. Le but de cet article n'est donc pas de tester des hypothèses issues de la littérature scientifique, mais d'examiner dans quelle mesure la recherche contemporaine peut expliquer, confirmer ou parfois nuancer des observations de terrain.
Au-delà du cas du cerf élaphe, cette approche cherche finalement à répondre à une question plus large : dans quelle mesure une lecture géographique du territoire peut-elle aider à anticiper la présence des grands mammifères et à guider plus efficacement la prospection naturaliste ?
Introduction
From direct observation to geographical landscape reading
When a naturalist discovers an unfamiliar territory, one question quickly arises: where should one look for animals? This issue is particularly important in wildlife photography. Time available in the field is often limited, while potentially suitable habitats may cover dozens of square kilometres. In such a context, direct observation alone is not always sufficient. It becomes necessary to develop an approach to landscape interpretation that allows the identification of the most promising areas even before the first sighting.
This approach lies at the intersection of geography and ecology. It consists of viewing a territory not as a simple juxtaposition of habitats, but as a spatial system structured by altitudinal gradients, ecological interfaces, movement corridors, refuge areas and connections with neighbouring landscapes. Animals thus become not only subjects of observation, but also indicators of the territorial functioning of the landscapes they occupy.
The reflections presented in this article emerged from several years of wildlife observation and photography conducted in different Swiss mountain regions. Over time, certain recurring patterns became apparent. Why do some pastures appear to be regularly used while others remain empty? Why do certain valleys function as movement corridors whereas others seem to concentrate animals over long periods? Why do landscapes that are geographically very different nevertheless contain spatial structures that appear to fulfil similar functions?
These questions gradually led to a shift in perspective. The objective was no longer simply to locate animals, but to understand the spatial processes that organise their presence. Mountain passes, ridgelines, forest-open habitat interfaces, undisturbed areas and altitudinal gradients gradually became as important as the animals themselves.
Red deer (Cervus elaphus) provide a particularly interesting model for exploring these questions. The species is now widely distributed throughout the Swiss mountain ranges, making it possible to observe populations in a wide variety of landscape contexts. At the same time, it remains sufficiently elusive and mobile to make observation challenging. More unpredictable than many other large herbivores commonly encountered in mountain environments, red deer encourage observers to move beyond simple visual searching and to consider the functioning of the territory as a whole.
The observations presented here were made before any specific literature review was undertaken. Only afterwards were they compared with recent findings from spatial ecology, GPS telemetry and landscape ecology. The purpose of this article is therefore not to test hypotheses derived from scientific literature, but rather to examine the extent to which contemporary ecological research can explain, support or sometimes challenge field observations.
Beyond the specific case of red deer, this approach ultimately seeks to answer a broader question: to what extent can a geographical reading of a territory help anticipate the presence of large mammals and guide wildlife prospecting more effectively?
Einleitung
Von der direkten Beobachtung zum geografischen Lesen der Landschaft
Wenn ein Naturbeobachter ein unbekanntes Gebiet betritt, stellt sich rasch eine Frage: Wo sollte man nach Tieren suchen? Diese Frage ist besonders in der Wildtierfotografie wichtig. Die verfügbare Zeit im Gelände ist oft begrenzt, während potenziell geeignete Lebensräume viele Quadratkilometer umfassen können. Direkte Beobachtung allein reicht daher nicht immer aus. Es braucht eine Lesart der Landschaft, die vielversprechende Bereiche bereits vor der ersten Sichtung erkennbar macht.
Dieser Ansatz liegt an der Schnittstelle von Geografie und Ökologie. Ein Gebiet wird nicht als einfache Aneinanderreihung von Habitaten verstanden, sondern als räumliches System aus Höhengradienten, ökologischen Übergängen, Bewegungskorridoren, Rückzugsräumen und Verbindungen zu benachbarten Landschaften. Tiere werden dadurch auch zu Indikatoren für das territoriale Funktionieren der Landschaften, die sie nutzen.
Die hier vorgestellten Überlegungen entstanden aus mehreren Jahren der Naturbeobachtung und Wildtierfotografie in verschiedenen Schweizer Gebirgsräumen. Mit der Zeit traten wiederkehrende Muster hervor: Manche Weiden werden regelmäßig genutzt, andere bleiben leer; manche Täler wirken als Bewegungsachsen, während andere Tiere längerfristig konzentrieren; und sehr unterschiedliche Landschaften weisen Strukturen auf, die vergleichbare Funktionen erfüllen.
Diese Fragen führten schrittweise zu einem Perspektivwechsel. Ziel war nicht mehr nur, Tiere zu lokalisieren, sondern die räumlichen Prozesse zu verstehen, die ihre Anwesenheit organisieren. Pässe, Gratlinien, Wald-Offenland-Übergänge, störungsarme Bereiche und Höhengradienten wurden ebenso wichtig wie die Tiere selbst.
Der Rothirsch (Cervus elaphus) eignet sich besonders gut, um diese Fragen zu untersuchen. Die Art ist in den Schweizer Gebirgsräumen heute weit verbreitet und lässt sich in unterschiedlichen Landschaftskontexten beobachten. Zugleich bleibt sie diskret und mobil genug, um die Beobachtung anspruchsvoll zu machen. Der Rothirsch zwingt den Beobachter, über die reine visuelle Suche hinauszugehen und das Gebiet als Ganzes zu verstehen.
Die hier vorgestellten Beobachtungen wurden vor einer spezifischen Literaturrecherche gemacht und erst anschließend mit Ergebnissen aus Raumökologie, GPS-Telemetrie und Landschaftsökologie verglichen. Ziel ist nicht, Literaturhypothesen zu prüfen, sondern zu zeigen, wie Forschungsergebnisse Felderfahrungen erklären, stützen oder nuancieren können.
Über den Rothirsch hinaus stellt sich damit eine allgemeinere Frage: In welchem Maß kann ein geografisches Lesen des Gebiets helfen, die Anwesenheit großer Säugetiere vorauszudenken und die Feldsuche gezielter zu führen?
Introduzione
Dall'osservazione diretta alla lettura geografica del paesaggio
Quando un naturalista scopre un territorio sconosciuto, emerge subito una domanda: dove cercare gli animali? Questa questione è particolarmente importante nella fotografia naturalistica. Il tempo disponibile sul campo è spesso limitato, mentre gli habitat potenzialmente favorevoli possono estendersi su decine di chilometri quadrati. In questo contesto l'osservazione diretta, da sola, non basta sempre. Diventa necessario sviluppare una lettura del paesaggio capace di individuare i settori più promettenti prima ancora del primo avvistamento.
Questo approccio appartiene tanto alla geografia quanto all'ecologia. Consiste nel considerare il territorio non come una semplice giustapposizione di habitat, ma come un sistema spaziale strutturato da gradienti altitudinali, interfacce ecologiche, corridoi di movimento, zone rifugio e connessioni con i paesaggi vicini. Gli animali diventano così anche indicatori del funzionamento territoriale dei paesaggi che occupano.
Le riflessioni presentate in questo articolo nascono da diversi anni di osservazione e fotografia faunistica in varie regioni montane svizzere. Nel tempo sono apparse alcune regolarità: alcuni pascoli sembrano frequentati regolarmente, altri restano vuoti; certe valli funzionano come assi di spostamento, mentre altre concentrano gli animali più a lungo; paesaggi molto diversi possono contenere strutture spaziali con funzioni simili.
Queste domande hanno portato progressivamente a un cambiamento di prospettiva. L'obiettivo non era più semplicemente localizzare gli animali, ma comprendere i processi spaziali che organizzano la loro presenza. Passi, creste, interfacce bosco-spazio aperto, aree poco disturbate e gradienti altitudinali sono diventati importanti quanto gli animali stessi.
Il cervo nobile (Cervus elaphus) è un modello particolarmente interessante per esplorare queste domande. Oggi la specie è ampiamente distribuita nei massicci svizzeri, il che permette di osservarla in contesti paesaggistici variati. Resta però abbastanza discreta e mobile da rendere l'osservazione esigente. Più imprevedibile di molti altri grandi erbivori montani, il cervo spinge l'osservatore a superare la semplice ricerca visiva e a interessarsi al funzionamento del territorio nel suo insieme.
Le osservazioni presentate qui sono state realizzate prima di una ricerca bibliografica specifica e solo in seguito confrontate con risultati di ecologia spaziale, telemetria GPS ed ecologia del paesaggio. Lo scopo non è verificare ipotesi tratte dalla letteratura, ma capire in che misura la ricerca scientifica possa illuminare, confermare o talvolta sfumare le osservazioni di campo.
Al di là del caso specifico del cervo, questo approccio cerca infine di rispondere a una domanda più ampia: fino a che punto una lettura geografica del territorio può aiutare ad anticipare la presenza dei grandi mammiferi e guidare più efficacemente la prospezione naturalistica?
1. Structure du paysage
La structure du paysage organise les déplacements
La première régularité apparue au fil des observations concerne l'importance de la connectivité. Dans plusieurs régions de montagne, certains secteurs semblaient fonctionner principalement comme des zones de transit plutôt que comme des sites d'occupation durable.
Le Val di Blenio, au Tessin, constitue un exemple particulièrement intéressant. Cette vaste vallée alpine orientée nord-sud relie le col du Lukmanier aux plaines tessinoises et forme un axe de déplacement naturel entre plusieurs systèmes montagneux. Des observations répétées suggèrent que certains secteurs y fonctionnent davantage comme des corridors que comme des habitats permanents : les animaux y apparaissent souvent brièvement avant de disparaître vers des vallées latérales ou des versants voisins.
Une logique comparable a été observée dans la Combe de l'A, en Valais. Cette vallée alpine est dominée par de vastes pentes herbeuses, des pierriers et des lignes de crête dépassant 2500 mètres d'altitude. Les cols et les passages d'altitude semblent y jouer un rôle d'interface entre plusieurs bassins versants au sein d'un système fonctionnel plus vaste. Les cerfs observés dans les pelouses alpines supérieures ne semblaient pas liés à une seule vallée, mais à un réseau territorial beaucoup plus large.
Des phénomènes comparables ont été documentés dans le Binntal, dans le Haut-Valais oriental. Cette vallée alpine de haute altitude est reliée à plusieurs vallées italiennes par des cols dépassant fréquemment 2400 à 2500 mètres. Des suivis réalisés dans la région montrent que certains cerfs franchissent régulièrement ces passages pour rejoindre des vallées méridionales durant l'hiver. Ces observations rappellent que les limites visibles du paysage correspondent rarement aux limites fonctionnelles réellement utilisées par les animaux.
À l'inverse, le Justistal, dans l'Oberland bernois, présente une organisation spatiale beaucoup plus contrainte. Cette vallée relativement étroite est bordée de versants abrupts et de massifs forestiers qui compartimentent fortement l'espace. Les déplacements semblent concentrés dans un nombre limité d'itinéraires préférentiels reliant différentes unités territoriales. Les animaux paraissent donc plus dépendants d'un réseau restreint de passages reliant les secteurs voisins. Urbina et al. (2023) ont montré que les déplacements du cerf en Suisse occidentale s'organisent autour de corridors fonctionnels reliant des noyaux d'habitat, certaines zones agissant comme de véritables goulets d'étranglement régionaux.
La région du Marchairuz, dans le Jura vaudois, illustre une autre situation. Le paysage y forme une large mosaïque de pâturages boisés, de forêts ouvertes et de clairières où la topographie impose beaucoup moins de contraintes. Les déplacements semblent moins dépendants de quelques corridors identifiables que de la continuité générale du paysage. La connectivité y repose davantage sur une mosaïque diffuse que sur des axes de déplacement clairement définis.
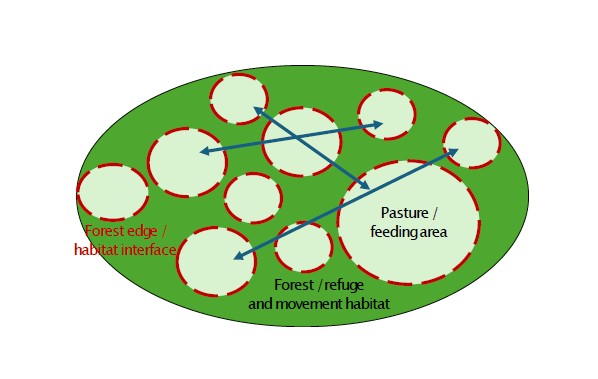
Une seconde régularité concerne le rôle des interfaces entre milieux ouverts et couverts. Dans l'ensemble des régions considérées, les zones de gagnage les plus utilisées partagent une caractéristique commune : la proximité immédiate d'un couvert permettant un repli rapide. Cette tendance est particulièrement visible au Marchairuz, où les animaux utilisent préférentiellement les lisières, les pâturages boisés et les clairières entourées de forêt. Elle se retrouve également dans les vallées alpines, où les espaces ouverts éloignés des refuges semblent beaucoup moins fréquentés. Les travaux de Rempfler et al. (2025) confirment cette tendance en montrant que les cerfs sélectionnent préférentiellement les secteurs proches du couvert forestier et évitent généralement les milieux ouverts trop éloignés d'un refuge potentiel.
Avant même de considérer les ressources alimentaires, le territoire apparaît donc déjà structuré par deux éléments fondamentaux : la connectivité et l'organisation des interfaces d'habitat. Ces observations suggèrent que les territoires utilisés par le cerf peuvent être regroupés en quelques grands modèles d'organisation spatiale.
1. Landscape structure
Landscape structure organises movements
The first recurring pattern that emerged from field observations concerns the importance of connectivity. In several mountain regions, certain areas appeared to function primarily as transit zones rather than as long-term occupation sites.
The Val di Blenio in Ticino provides a particularly interesting example. This large north-south oriented Alpine valley links the Lukmanier Pass to the Ticino lowlands and forms a natural movement axis between several mountain systems. Repeated observations suggested that some sectors functioned more as corridors than as permanent habitat. Animals frequently appeared only briefly before disappearing towards side valleys or neighbouring slopes.
A similar pattern was observed in the Combe de l'A, in the canton of Valais. This Alpine valley is dominated by extensive grass slopes, scree fields and ridgelines rising above 2,500 metres. High-altitude passes and crossing points appear to function as interfaces connecting several drainage basins within a larger functional system. Red deer observed in the upper alpine meadows did not seem to be associated with a single valley but rather with a much broader territorial network.
Comparable phenomena have been documented in the Binntal, in eastern Upper Valais. This high-altitude Alpine valley is connected to several Italian valleys through passes frequently exceeding 2,400-2,500 metres. Monitoring studies conducted in the region have shown that some deer regularly cross these passes in order to reach southern valleys during winter. Such observations remind us that visible landscape boundaries rarely correspond to the functional boundaries actually used by animals.
By contrast, the Justistal in the Bernese Oberland displays a much more constrained spatial organisation. This relatively narrow valley is bordered by steep slopes and forested massifs that strongly compartmentalise space. Movements appear concentrated within a limited number of preferred routes connecting different territorial units. Animals therefore seem more dependent on a restricted network of passages linking neighbouring areas. Urbina et al. (2023) showed that red deer movements in western Switzerland are organised around functional corridors connecting habitat cores. Certain areas act as genuine bottlenecks where movements become concentrated at the regional scale.
The Marchairuz region in the Vaud Jura illustrates yet another situation. The landscape consists of a broad mosaic of wooded pastures, open forests and clearings where topography imposes far fewer constraints. Movements appear less dependent on a few identifiable corridors than on the overall continuity of the landscape. Connectivity here is provided by a diffuse mosaic rather than by clearly defined movement axes.
Recent GPS telemetry studies provide valuable support for these observations.
A second recurring pattern concerns the role of interfaces between open and covered habitats. Across all the mountain regions considered, the most frequently used feeding areas shared one common characteristic: immediate proximity to cover allowing rapid retreat. This pattern is particularly evident in the Marchairuz region, where animals preferentially use forest edges, wooded pastures and clearings surrounded by woodland. It is also visible in Alpine valleys, where open areas located far from refuge habitats appear to be used much less frequently. The work of Rempfler et al. (2025) confirms this tendency, showing that red deer preferentially select areas located close to forest cover and generally avoid open habitats situated too far from potential refuges.
Even before food resources are considered, the territory already appears to be structured by two fundamental elements: connectivity and the organisation of habitat interfaces. These observations suggest that territories used by red deer can be grouped into a limited number of broad spatial organisation models.
1. Landschaftsstruktur
Die Landschaftsstruktur organisiert Bewegungen
Das erste wiederkehrende Muster betrifft die Bedeutung der Vernetzung. In mehreren Gebirgsräumen schienen bestimmte Bereiche vor allem als Transitflächen zu dienen, nicht als dauerhaft genutzte Aufenthaltsräume.
Das Val di Blenio im Tessin ist dafür ein besonders interessantes Beispiel. Dieses große, nord-südlich ausgerichtete Alpental verbindet den Lukmanierpass mit den Tessiner Niederungen und bildet eine natürliche Bewegungsachse zwischen mehreren Gebirgssystemen. Wiederholte Beobachtungen deuten darauf hin, dass einige Sektoren eher als Korridore denn als Dauerhabitat funktionieren: Tiere erscheinen dort oft nur kurz, bevor sie in Seitentäler oder auf benachbarte Hänge verschwinden.
Eine ähnliche Logik wurde in der Combe de l'A im Wallis beobachtet. Diese alpine Talstruktur ist geprägt von weiten Grasflanken, Schutthalden und Graten über 2500 Metern. Hoch gelegene Pässe und Übergänge verbinden mehrere Einzugsgebiete innerhalb eines größeren funktionalen Systems. Rothirsche in den oberen alpinen Matten wirkten nicht an ein einzelnes Tal gebunden, sondern an ein viel größeres territoriales Netzwerk.
Ähnliche Phänomene sind im Binntal im östlichen Oberwallis dokumentiert. Dieses hoch gelegene Alpental ist über Pässe von häufig über 2400 bis 2500 Metern mit italienischen Tälern verbunden. Untersuchungen zeigen, dass manche Hirsche diese Übergänge regelmäßig nutzen, um im Winter südliche Täler zu erreichen. Sichtbare Landschaftsgrenzen entsprechen also selten den funktionalen Grenzen, die Tiere tatsächlich nutzen.
Im Gegensatz dazu zeigt das Justistal im Berner Oberland eine deutlich stärker eingeschränkte räumliche Organisation. Das enge Tal ist von steilen Hängen und Waldmassiven geprägt, die den Raum stark gliedern. Bewegungen konzentrieren sich auf wenige bevorzugte Passagen zwischen verschiedenen Gebietseinheiten. Urbina et al. (2023) zeigen, dass Bewegungen des Rothirsches in der Westschweiz um funktionale Korridore organisiert sind, die Habitatkerne verbinden. Einige Bereiche wirken als regionale Engstellen.
Der Marchairuz im Waadtländer Jura zeigt wiederum eine andere Situation. Dort bilden Waldweiden, offene Wälder und Lichtungen eine breite Mosaiklandschaft, in der das Relief weniger einschränkend wirkt. Die Bewegungen hängen weniger von einzelnen klaren Korridoren ab als von der allgemeinen Kontinuität der Landschaft.
Ein zweites Muster betrifft Übergänge zwischen offenen und gedeckten Lebensräumen. Die regelmäßig genutzten Äsungsflächen liegen fast immer nahe an Deckung, die einen raschen Rückzug ermöglicht. Rempfler et al. (2025) bestätigen diese Tendenz: Rothirsche wählen bevorzugt Bereiche in der Nähe von Walddeckung und meiden offene Flächen, die zu weit von Rückzugsmöglichkeiten entfernt sind.
Schon bevor Nahrungsressourcen betrachtet werden, erscheint das Gebiet durch zwei grundlegende Elemente strukturiert: Vernetzung und Habitatübergänge. Daraus ergibt sich die Möglichkeit, Rothirschgebiete in einige größere räumliche Organisationsmodelle einzuordnen.
1. Struttura del paesaggio
La struttura del paesaggio organizza gli spostamenti
La prima regolarità emersa dalle osservazioni riguarda l'importanza della connettività. In diverse regioni montane, alcuni settori sembravano funzionare soprattutto come zone di transito più che come aree di occupazione duratura.
Il Val di Blenio, in Ticino, offre un esempio particolarmente interessante. Questa ampia valle alpina orientata nord-sud collega il passo del Lucomagno alle pianure ticinesi e costituisce un asse naturale di movimento tra più sistemi montuosi. Osservazioni ripetute suggeriscono che alcuni settori funzionino più come corridoi che come habitat permanenti: gli animali vi appaiono spesso brevemente prima di scomparire verso valli laterali o versanti vicini.
Una logica simile è stata osservata nella Combe de l'A, in Vallese. Questa valle alpina è dominata da ampi versanti erbosi, ghiaioni e creste oltre i 2500 metri. I passi e i punti di attraversamento in quota sembrano funzionare come interfacce tra più bacini versanti all'interno di un sistema funzionale più ampio. I cervi osservati nei pascoli alpini superiori non sembravano legati a una sola valle, ma a una rete territoriale molto più ampia.
Fenomeni analoghi sono documentati nel Binntal, nell'Alto Vallese orientale. Questa valle alpina di alta quota è collegata a più valli italiane attraverso passi che superano spesso i 2400-2500 metri. Alcuni cervi attraversano regolarmente questi passaggi per raggiungere le valli meridionali durante l'inverno. Queste osservazioni ricordano che i limiti visibili del paesaggio raramente coincidono con i limiti funzionali realmente utilizzati dagli animali.
Al contrario, il Justistal, nell'Oberland bernese, presenta un'organizzazione molto più vincolata. La valle, relativamente stretta, è bordata da versanti ripidi e massicci forestali che compartimentano fortemente lo spazio. Gli spostamenti si concentrano in pochi passaggi preferenziali. Urbina et al. (2023) mostrano che gli spostamenti del cervo nella Svizzera occidentale si organizzano attorno a corridoi funzionali che collegano nuclei di habitat, con veri colli di bottiglia a scala regionale.
Il Marchairuz, nel Giura vodese, illustra ancora un'altra situazione. Il paesaggio è formato da un mosaico di pascoli arborati, foreste aperte e radure, dove il rilievo impone meno vincoli. Gli spostamenti dipendono meno da corridoi chiaramente identificabili che dalla continuità generale del paesaggio.
Una seconda regolarità riguarda il ruolo delle interfacce tra ambienti aperti e coperti. In tutti i massicci osservati, le aree di alimentazione più regolarmente frequentate presentano quasi sempre una caratteristica comune: la vicinanza immediata a un riparo che consente un rapido ritiro. Rempfler et al. (2025) confermano questa tendenza, mostrando che i cervi selezionano preferenzialmente i settori vicini alla copertura forestale ed evitano gli spazi aperti troppo lontani da un possibile rifugio.
Prima ancora di considerare le risorse alimentari, il territorio appare dunque strutturato da due elementi fondamentali: connettività e organizzazione delle interfacce di habitat. Queste osservazioni permettono di raggruppare i territori utilizzati dal cervo in alcuni grandi modelli di organizzazione spaziale.
2. Typologie territoriale
Une typologie de l'organisation territoriale
La diversité des territoires examinés soulève une question plus générale. Au-delà des caractéristiques propres à chaque région de montagne, existe-t-il des formes récurrentes d'organisation spatiale permettant de comparer des paysages très différents ? Il ne s'agit pas de réduire la complexité des territoires réels à quelques catégories rigides. Chaque massif possède son histoire, sa topographie, ses usages humains et ses dynamiques écologiques.
La valeur d'un tel modèle est surtout heuristique. Il ne cherche pas à décrire chaque territoire de manière exhaustive, mais à fournir un cadre simplifié pour identifier les principaux facteurs qui structurent les déplacements et l'utilisation de l'espace par le cerf. Cette approche facilite les comparaisons entre régions, met en évidence les éléments fonctionnels des paysages et constitue une première étape vers une lecture prédictive du territoire.
Sans prétendre proposer une typologie exhaustive, les observations présentées ici suggèrent l'existence de trois grands modèles territoriaux. Ces modèles ne sont pas exclusifs : un même massif peut combiner localement plusieurs logiques spatiales. Ils offrent néanmoins un cadre utile pour comprendre comment les cerfs organisent leur espace.
2. Territorial typology
A typology of territorial organisation
The diversity of the territories examined raises a broader question. Beyond the specific characteristics of each mountain region, are there recurring forms of spatial organisation that make it possible to compare very different landscapes? The objective is not, of course, to reduce the complexity of real territories to a few rigid categories. Each massif has its own history, topography, human land use and ecological dynamics. Nevertheless, comparing observations made in different contexts and confronting them with findings from the scientific literature reveals a number of recurring spatial configurations.
The value of such a model is primarily heuristic. It does not seek to describe every territory faithfully, but rather to provide a simplified framework for identifying the principal factors that structure movements and space use by red deer. This approach facilitates comparisons between regions, highlights the functional elements of landscapes and provides a first step towards a predictive reading of territory.
Without claiming to offer an exhaustive typology, the observations presented here suggest the existence of three broad territorial models. These models are not mutually exclusive. A single mountain region may locally combine several spatial logics. Nevertheless, they provide a useful framework for understanding how deer organise their space and for comparing territories with very different physical characteristics.
2. Territoriale Typologie
Eine Typologie territorialer Organisation
Die Vielfalt der untersuchten Gebiete führt zu einer allgemeinen Frage: Gibt es wiederkehrende Formen räumlicher Organisation, die einen Vergleich sehr unterschiedlicher Landschaften ermöglichen? Ziel ist nicht, die Komplexität realer Gebiete auf starre Kategorien zu reduzieren. Jedes Massiv hat seine eigene Geschichte, Topografie, Landnutzung und ökologische Dynamik.
Der Wert eines solchen Modells ist vor allem heuristisch. Es beschreibt nicht jedes Gebiet vollständig, sondern liefert einen vereinfachten Rahmen, um die wichtigsten Faktoren zu erkennen, die Bewegungen und Raumnutzung des Rothirsches strukturieren. Dadurch werden regionale Vergleiche erleichtert und funktionale Landschaftselemente hervorgehoben.
Die Beobachtungen deuten auf drei große territoriale Modelle hin. Sie schließen sich nicht gegenseitig aus; ein einzelnes Gebirge kann lokal mehrere räumliche Logiken verbinden. Dennoch bieten sie einen nützlichen Rahmen, um die Organisation des Rothirschraums zu verstehen.
2. Tipologia territoriale
Una tipologia dell'organizzazione territoriale
La diversità dei territori esaminati conduce a una domanda più generale: esistono forme ricorrenti di organizzazione spaziale che permettono di confrontare paesaggi molto diversi? L'obiettivo non è ridurre la complessità dei territori reali a poche categorie rigide. Ogni massiccio possiede una propria storia, topografia, utilizzazione umana e dinamica ecologica.
Il valore di questo modello è soprattutto euristico. Non mira a descrivere ogni territorio in modo completo, ma offre una griglia semplificata per individuare i principali fattori che strutturano gli spostamenti e l'uso dello spazio da parte del cervo. Questo approccio facilita il confronto tra regioni e mette in evidenza gli elementi funzionali dei paesaggi.
Le osservazioni suggeriscono l'esistenza di tre grandi modelli territoriali. Questi modelli non sono esclusivi: uno stesso massiccio può combinare localmente più logiche spaziali. Tuttavia offrono un quadro utile per comprendere come i cervi organizzano il loro spazio.
A. Système continu en mosaïque
A. Continuous mosaic system
A. Kontinuierliches Mosaiksystem
A. Sistema continuo a mosaico
Les ressources alimentaires, les zones de repos et les interfaces forêt-milieu ouvert sont réparties dans un paysage relativement continu. Les déplacements sont faiblement contraints et dépendent surtout de la continuité générale de l'environnement. Ce modèle caractérise les régions de moyenne montagne combinant forêts et paysages pastoraux, comme le Jura vaudois. Exemple : le Marchairuz, où pâturages boisés, clairières et forêts ouvertes forment un réseau continu favorable aux déplacements de la faune.
Food resources, resting areas and forest-open habitat interfaces are distributed within a relatively continuous landscape. Movements are only weakly constrained and depend primarily on the overall continuity of the environment. This model is characteristic of mid-altitude mountain regions combining forests and pastoral landscapes, such as the Vaud Jura. Example: the Marchairuz region, where wooded pastures, clearings and open forests form a continuous network favourable to wildlife movements.
Nahrungsflächen, Ruhebereiche und Wald-Offenland-Übergänge liegen in einer relativ kontinuierlichen Landschaft. Bewegungen sind nur schwach eingeschränkt und beruhen vor allem auf der allgemeinen Kontinuität des Raums. Dieses Modell ist typisch für Mittelgebirgsräume mit Wald- und Weidelandschaften, etwa den Waadtländer Jura. Beispiel: Marchairuz, wo Waldweiden, Lichtungen und offene Wälder ein zusammenhängendes Netzwerk bilden.
Risorse alimentari, aree di riposo e interfacce bosco-spazio aperto sono distribuite in un paesaggio relativamente continuo. Gli spostamenti sono poco vincolati e dipendono soprattutto dalla continuità generale dell'ambiente. Questo modello è tipico delle regioni di media montagna che combinano foreste e paesaggi pastorali, come il Giura vodese. Esempio: il Marchairuz, dove pascoli arborati, radure e foreste aperte formano una rete continua favorevole ai movimenti della fauna.
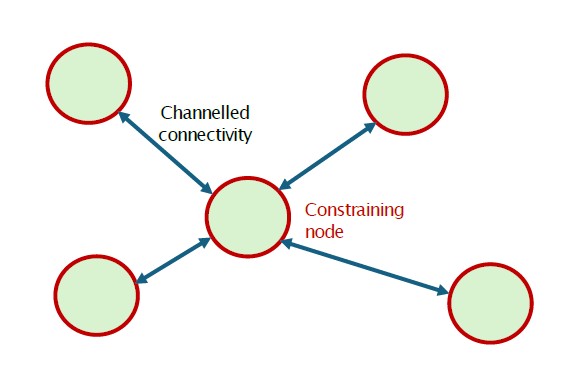
B. Système fragmenté à connectivité contrainte
B. Fragmented system with constrained connectivity
B. Fragmentiertes System mit eingeschränkter Vernetzung
B. Sistema frammentato a connettività vincolata
Les habitats favorables sont répartis entre des noyaux distincts reliés par un nombre limité de corridors ou de points de passage. Les déplacements se concentrent le long d'itinéraires fonctionnels qui maintiennent la connectivité du système. Cette organisation apparaît dans les vallées fortement compartimentées comme dans les paysages humanisés où la fragmentation canalise les mouvements. Exemples : le Justistal, où les passages entre massifs forestiers sont peu nombreux, et le Piano di Magadino, où les habitats favorables restent connectés dans un paysage fortement modifié.
Suitable habitats are distributed among distinct core areas connected by a limited number of corridors or crossing points. Movements become concentrated along functional routes that maintain connectivity within the system. This organisation occurs both in strongly compartmentalised valleys and in human-dominated landscapes where habitat fragmentation channels wildlife movements. Refuge areas and corridors become the key structural elements of the territory. Examples: the Justistal, where movements are concentrated through a small number of passages between forest massifs, and the Piano di Magadino, where suitable habitats remain connected across a heavily modified landscape.
Geeignete Habitate liegen in getrennten Kernbereichen, die durch wenige Korridore oder Übergänge verbunden sind. Bewegungen konzentrieren sich auf funktionale Routen, die die Vernetzung des Systems aufrechterhalten. Diese Organisation tritt in stark gegliederten Tälern ebenso auf wie in vom Menschen geprägten Landschaften, in denen Fragmentierung die Bewegungen kanalisiert. Beispiele: Justistal und Piano di Magadino.
Gli habitat favorevoli sono distribuiti in nuclei distinti collegati da un numero limitato di corridoi o punti di passaggio. Gli spostamenti si concentrano lungo rotte funzionali che mantengono la connettività del sistema. Questa organizzazione si trova sia in valli fortemente compartimentate sia in paesaggi dominati dall'uomo, dove la frammentazione canalizza i movimenti della fauna. Esempi: Justistal e Piano di Magadino.
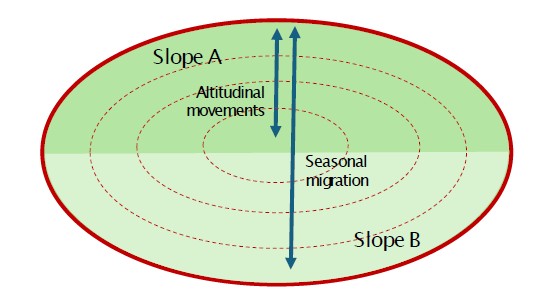
C. Système altitudinal multiversant
C. Multi-slope altitudinal system
C. Höhenbezogenes Mehrhangsystem
C. Sistema altitudinale multiversante
L'organisation spatiale repose sur des déplacements saisonniers entre différents étages altitudinaux et plusieurs versants appartenant à une même unité territoriale. Les cols et les lignes de crête relient les secteurs utilisés au cours de l'année. Ce modèle est fréquent dans les vallées alpines où l'enneigement et la disponibilité des ressources génèrent une forte mobilité saisonnière. Exemples : la Combe de l'A et le Binntal.
Spatial organisation is based on seasonal movements between different altitudinal zones and multiple slopes belonging to the same territorial unit. Mountain passes and ridgelines provide connections between the sectors used throughout the year. This model is common in Alpine valleys where snow conditions and resource availability generate strong seasonal mobility. Examples: the Combe de l'A and the Binntal, where red deer successively exploit several altitudinal levels and regularly cross passes and ridgelines in order to move from one slope to another.
Die räumliche Organisation beruht auf jahreszeitlichen Bewegungen zwischen verschiedenen Höhenstufen und mehreren Hängen derselben Gebietseinheit. Pässe und Gratlinien verbinden die im Jahresverlauf genutzten Sektoren. Dieses Modell ist in Alpentalräumen häufig, wo Schneelage und Ressourcenverfügbarkeit starke saisonale Mobilität erzeugen. Beispiele: Combe de l'A und Binntal.
L'organizzazione spaziale si basa su spostamenti stagionali tra diversi piani altitudinali e più versanti appartenenti alla stessa unità territoriale. Colli e linee di cresta collegano i settori utilizzati durante l'anno. Questo modello è comune nelle valli alpine dove neve e disponibilità di risorse generano una forte mobilità stagionale. Esempi: Combe de l'A e Binntal.
3. Saisons, climat et météo
Les saisons, le climat et la météo redistribuent l'utilisation de l'espace
Si le paysage définit les possibilités disponibles pour les animaux, la saison détermine lesquelles de ces possibilités seront effectivement utilisées.
L'hiver correspond à une période de mobilité relativement limitée. L'enneigement augmente le coût énergétique des déplacements et réduit fortement l'espace réellement disponible. Les populations tendent donc à se concentrer dans les secteurs les moins contraignants. Les comptages réalisés dans l'Intyamon, en Haute-Gruyère, et dans le Pays-d'Enhaut voisin illustrent bien ce phénomène : lors de fortes chutes de neige, plusieurs centaines d'animaux peuvent descendre vers les fonds de vallée autour de 700 mètres d'altitude avant de recoloniser progressivement les altitudes supérieures au printemps.
Le Piano di Magadino, au Tessin, offre un autre exemple. Cette grande plaine alluviale située entre Bellinzone et Locarno est entourée d'infrastructures de transport, d'espaces urbains et d'agriculture intensive. Malgré ce contexte très modifié, la réserve naturelle des Bolle di Magadino accueille une importante population hivernante de cerfs. Le climat plus doux, l'enneigement limité et la présence de refuges de qualité compensent en partie les contraintes humaines. Lorsque les températures remontent et que les ressources redeviennent disponibles dans les montagnes voisines, les animaux se dispersent progressivement vers les vallées et les hauteurs environnantes.
Le printemps est probablement la période la plus dynamique de l'année. Les observations menées dans le Val di Blenio suggéraient que les animaux suivaient étroitement la fonte des neiges et l'émergence de la nouvelle végétation. Les versants exposés au sud devenaient attractifs plusieurs semaines avant les secteurs voisins. Les travaux de Sigrist et al. (2022) soutiennent cette interprétation en montrant que les cerfs suivent de près la progression du green-up, c'est-à-dire l'apparition de la végétation fraîche.
L'été correspond généralement à une phase plus stable. Les ressources deviennent abondantes et les animaux s'appuient davantage sur des habitats locaux. Les grands déplacements liés au suivi du green-up diminuent progressivement et sont remplacés par une utilisation plus régulière des territoires estivaux. Dans les régions alpines, les cerfs peuvent alors occuper des altitudes plus élevées et élargir le gradient altitudinal exploité. Dans la Combe de l'A, les grands mâles utilisent régulièrement les pelouses alpines et les milieux minéraux situés au-dessus de 2200 mètres.
L'automne représente une phase particulière du cycle annuel. Les déplacements associés au brame peuvent temporairement reconnecter des territoires éloignés. Plusieurs études menées en Suisse ont documenté des déplacements de plusieurs dizaines de kilomètres reliant différents massifs. Certaines populations forment de véritables réseaux en étoile centrés sur des places de brame majeures et impliquant des animaux venus de régions éloignées.
La synthèse saisonnière proposée ici ne provient pas d'une seule étude. Elle résulte de la comparaison d'observations menées dans plusieurs régions de montagne et de résultats convergents de la littérature scientifique. Les études de télémétrie GPS montrent notamment que les facteurs qui structurent l'utilisation de l'espace varient fortement au cours de l'année.
3. Seasons, climate and weather
Seasons, climate and weather redistribute the use of space
If the landscape defines the possibilities available to animals, the season determines which of these possibilities will actually be used.
Winter corresponds to a period of relatively limited mobility. Snow cover increases the energetic cost of movement and greatly reduces the amount of space that is effectively available. Populations therefore tend to concentrate in the least restrictive sectors. Counts conducted in the Intyamon region of Haute-Gruyère and in the neighbouring Pays-d'Enhaut illustrate this phenomenon particularly well. This area forms a large ecological unit linking the Fribourg and Vaud Pre-Alps and supports one of the highest red deer densities in western Switzerland. During periods of heavy snowfall, several hundred animals may descend to valley bottoms around 700 metres above sea level before progressively recolonising higher elevations in spring.
The Piano di Magadino in Ticino provides another interesting example. Located between Bellinzona and Locarno, this large alluvial plain is surrounded by transport infrastructure, urban areas and intensive agriculture. Despite this highly modified environment, the Bolle di Magadino nature reserve hosts an important wintering population of red deer. The milder climate, limited snow cover and availability of high-quality refuge habitats largely compensate for the constraints associated with human activities. As temperatures rise and resources become available again in the surrounding mountains, animals progressively disperse towards neighbouring valleys and uplands.
Spring is probably the most dynamic period of the year. Observations conducted in the Val di Blenio suggested that animals closely followed snowmelt and the emergence of new vegetation. South-facing slopes became attractive several weeks before neighbouring sectors. The work of Sigrist et al. (2022) supports this interpretation by showing that red deer closely track the progression of green-up, that is, the appearance of newly emerging vegetation. This period also appears to correspond to the phase of greatest annual mobility, during which altitudinal and sometimes regional movements become particularly important.
Summer generally corresponds to a more stable phase. Resources become abundant and animals rely more heavily on local habitats. The large-scale movements associated with tracking green-up progressively decline and are replaced by a more regular use of summer territories. In Alpine regions, red deer may then occupy higher elevations and expand the altitudinal range they exploit. In the Combe de l'A, large males regularly use alpine grasslands and mineral habitats situated above 2,200 metres. Animals nevertheless continue to seek a balance between resource quality, tranquillity and proximity to refuge areas. Movements remain significant but take place within a relatively stable summer territory.
Autumn represents a particular phase of the annual cycle. Movements associated with the rut may temporarily reconnect territories located far apart. Several studies conducted in Switzerland have documented movements of several tens of kilometres linking different mountain ranges. Some populations form genuine star-shaped networks centred on major rutting areas and involving animals originating from distant regions. These processes, however, are specific to the rutting period and have not been considered further in the present analysis.
The seasonal synthesis proposed here is not derived from a single study. Rather, it results from the comparison of observations conducted across several mountain regions and a number of convergent findings from the scientific literature. GPS telemetry studies notably demonstrate that the factors structuring space use vary considerably throughout the year. Snow cover and accessibility dominate during winter, green-up and connectivity become central in spring, while resource exploitation and the balance between feeding opportunities and safety play a greater role during summer. Autumn is characterised by rut-related movements and the temporary reconnection of territories that may otherwise remain largely separate.
3. Jahreszeiten, Klima und Wetter
Jahreszeiten, Klima und Wetter verteilen die Raumnutzung neu
Wenn die Landschaft die Möglichkeiten vorgibt, entscheidet die Jahreszeit, welche davon tatsächlich genutzt werden.
Der Winter ist eine Phase relativ eingeschränkter Mobilität. Schnee erhöht die energetischen Kosten der Fortbewegung und reduziert den effektiv verfügbaren Raum. Populationen konzentrieren sich daher in den am wenigsten einschränkenden Bereichen. Zählungen in der Intyamon-Region der Haute-Gruyère und im benachbarten Pays-d'Enhaut zeigen dieses Phänomen deutlich: Bei starkem Schneefall können mehrere hundert Tiere in Talböden um 700 Meter absteigen und im Frühling höhere Lagen schrittweise wieder besiedeln.
Das Piano di Magadino im Tessin ist ein weiteres Beispiel. Diese große Schwemmebene zwischen Bellinzona und Locarno ist von Verkehrsachsen, Siedlungsflächen und intensiver Landwirtschaft umgeben. Trotz dieser stark veränderten Umgebung beherbergt das Naturschutzgebiet Bolle di Magadino eine bedeutende überwinternde Rothirschpopulation. Mildes Klima, geringe Schneedecke und hochwertige Rückzugsräume kompensieren einen Teil der menschlichen Einschränkungen.
Der Frühling ist vermutlich die dynamischste Phase des Jahres. Beobachtungen im Val di Blenio deuten darauf hin, dass die Tiere der Schneeschmelze und dem frischen Pflanzenwachstum folgen. Südexponierte Hänge werden oft mehrere Wochen früher attraktiv als benachbarte Bereiche. Sigrist et al. (2022) stützen diese Interpretation, indem sie zeigen, dass Rothirsche das Green-up eng verfolgen.
Der Sommer entspricht meist einer stabileren Phase. Ressourcen sind reichlich vorhanden, und die Tiere nutzen lokale Habitate intensiver. In alpinen Räumen können Rothirsche höhere Lagen besetzen und den genutzten Höhengradienten erweitern. In der Combe de l'A nutzen starke Hirsche regelmäßig alpine Rasen und mineralische Lebensräume über 2200 Metern. Gleichzeitig bleibt das Gleichgewicht zwischen Ressourcenqualität, Ruhe und Rückzugsmöglichkeiten entscheidend.
Der Herbst ist eine besondere Phase des Jahreszyklus. Bewegungen im Zusammenhang mit der Brunft können weit entfernte Gebiete vorübergehend wieder verbinden. In der Schweiz wurden Bewegungen von mehreren Dutzend Kilometern dokumentiert, die verschiedene Gebirge miteinander verknüpfen.
Die hier vorgeschlagene jahreszeitliche Synthese stammt nicht aus einer einzelnen Studie. Sie ergibt sich aus dem Vergleich von Beobachtungen in mehreren Gebirgsräumen und konvergierenden Ergebnissen der Literatur. GPS-Telemetrie zeigt, dass die Faktoren der Raumnutzung im Jahresverlauf stark wechseln: Schnee und Zugänglichkeit im Winter, Green-up und Vernetzung im Frühling, Ressourcen und Sicherheit im Sommer sowie Brunftbewegungen im Herbst.
3. Stagioni, clima e meteorologia
Stagioni, clima e tempo ridistribuiscono l'uso dello spazio
Se il paesaggio definisce le possibilità disponibili per gli animali, la stagione determina quali di queste possibilità verranno effettivamente utilizzate.
L'inverno corrisponde a una fase di mobilità relativamente limitata. La neve aumenta il costo energetico degli spostamenti e riduce fortemente lo spazio realmente disponibile. Le popolazioni tendono quindi a concentrarsi nei settori meno vincolanti. I conteggi nell'Intyamon, in Haute-Gruyère, e nel vicino Pays-d'Enhaut illustrano bene questo fenomeno: durante forti nevicate, diverse centinaia di animali possono scendere verso i fondovalle intorno ai 700 metri prima di ricolonizzare progressivamente le quote superiori in primavera.
Il Piano di Magadino, in Ticino, offre un altro esempio. Questa grande pianura alluvionale tra Bellinzona e Locarno è circondata da infrastrutture di trasporto, aree urbane e agricoltura intensiva. Nonostante questo ambiente molto modificato, la riserva naturale delle Bolle di Magadino ospita un'importante popolazione svernante di cervi. Il clima più mite, la copertura nevosa limitata e la presenza di rifugi di qualità compensano in parte i vincoli umani.
La primavera è probabilmente il periodo più dinamico dell'anno. Le osservazioni nel Val di Blenio suggeriscono che gli animali seguano da vicino il disgelo e l'emergere della nuova vegetazione. I versanti esposti a sud diventano attrattivi diverse settimane prima dei settori vicini. Il lavoro di Sigrist et al. (2022) conferma questa interpretazione, mostrando che i cervi seguono la progressione del green-up.
L'estate corrisponde generalmente a una fase più stabile. Le risorse diventano abbondanti e gli animali si appoggiano maggiormente agli habitat locali. Nelle regioni alpine i cervi possono occupare quote più elevate e ampliare il gradiente altitudinale sfruttato. Nella Combe de l'A, i grandi maschi utilizzano regolarmente praterie alpine e ambienti minerali sopra i 2200 metri. Rimane però essenziale l'equilibrio tra qualità delle risorse, tranquillità e vicinanza ai rifugi.
L'autunno rappresenta una fase particolare del ciclo annuale. Gli spostamenti legati al bramito possono riconnettere temporaneamente territori lontani. Alcuni studi in Svizzera hanno documentato movimenti di diverse decine di chilometri tra massicci diversi.
La sintesi stagionale proposta qui non deriva da un singolo studio, ma dal confronto tra osservazioni condotte in varie regioni montane e risultati convergenti della letteratura. La telemetria GPS mostra che i fattori che strutturano l'uso dello spazio variano notevolmente durante l'anno: neve e accessibilità in inverno, green-up e connettività in primavera, risorse e sicurezza in estate, movimenti del bramito in autunno.
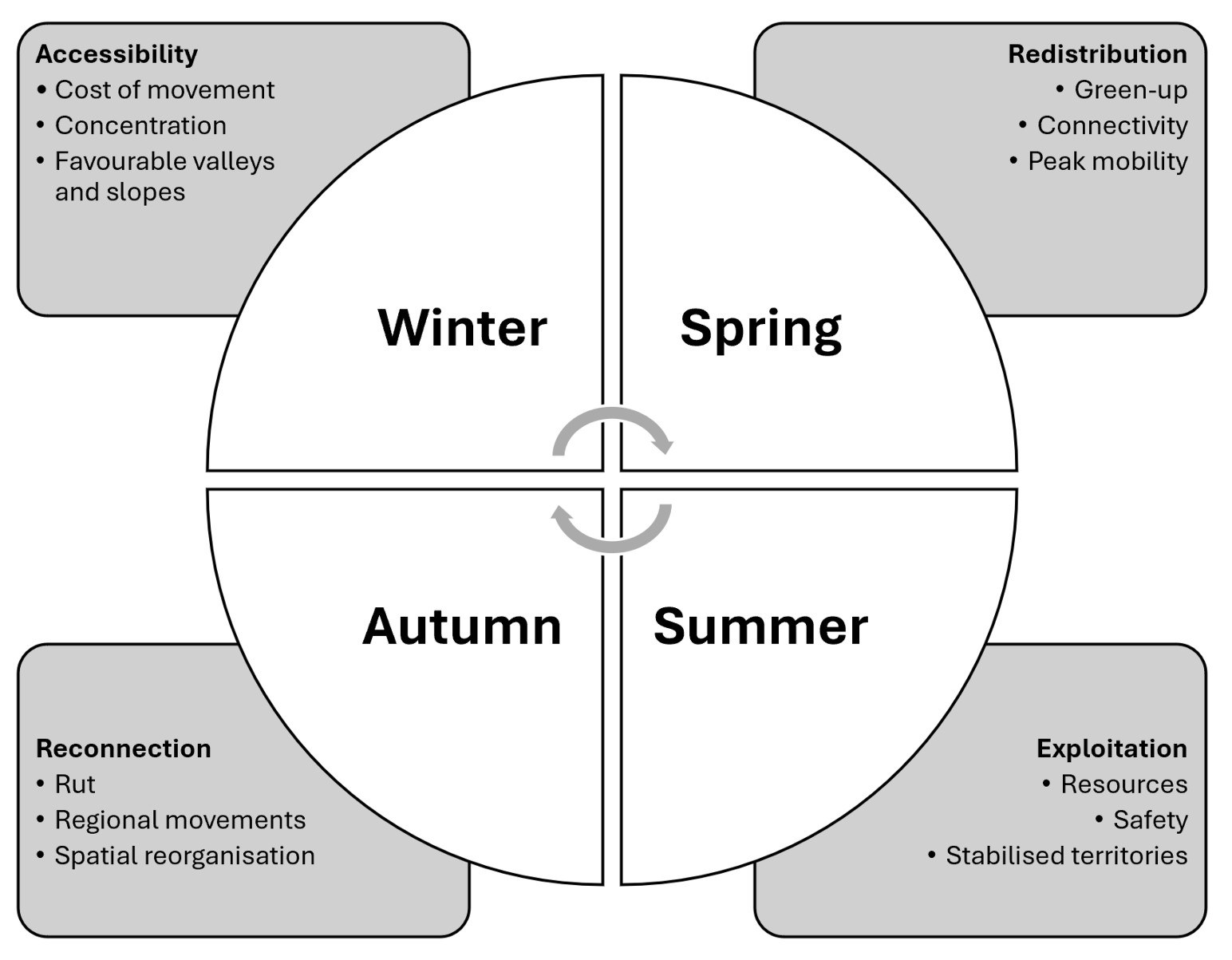
| Saison | Facteur dominant | Comportement spatial principal | Structures paysagères clés | Implications pour la prospection |
|---|---|---|---|---|
| Season | Dominant factor | Main spatial behaviour | Key landscape features | Implications for field prospecting |
| Jahreszeit | Dominanter Faktor | Räumliches Hauptverhalten | Wichtige Landschaftsmerkmale | Hinweise für die Feldsuche |
| Stagione | Fattore dominante | Comportamento spaziale principale | Caratteristiche chiave del paesaggio | Indicazioni per la prospezione |
| Hiver | Accessibilité | Déplacements réduits ; concentration dans les secteurs les moins coûteux sur le plan énergétique. | Fonds de vallée, versants sud, secteurs peu enneigés, lisières basses. | Se concentrer sur les zones accessibles, abritées, peu enneigées et proches d'un couvert. |
| Printemps | Connectivité + ressources | Forte mobilité ; suivi de la fonte des neiges et du green-up ; déplacements altitudinaux et parfois régionaux. | Versants sud, corridors, cols, vallées, interfaces forêt-pâturage. | Identifier les premières zones déneigées connectées à des habitats refuges. |
| Été | Ressources + sécurité | Stabilisation relative ; utilisation locale des zones de gagnage. | Pelouses alpines, pâturages, lisières, pierriers, zones de repos proches. | Rechercher le meilleur compromis entre fourrage de qualité, possibilités de fuite rapide et faible dérangement. |
| Automne | Connectivité territoriale | Réorganisation temporaire liée au brame ; déplacements parfois à longue distance. | Places de brame, vallées de convergence, cols, corridors de déplacement. | Identifier les zones de rassemblement et les corridors reliant plusieurs secteurs. |
| Winter | Accessibility | Reduced movements; concentration in the least energetically costly areas. | Valley bottoms, south-facing slopes, low-snow sectors, lower forest edges. | Focus on accessible, sheltered and lightly snow-covered areas close to cover. |
| Spring | Connectivity + resources | High mobility; tracking snowmelt and green-up; altitudinal and regional movements. | South-facing slopes, corridors, mountain passes, valleys, forest-pasture interfaces. | Identify the first snow-free areas connected to refuge habitats. |
| Summer | Resources + safety | Relative stabilisation; local use of feeding areas. | Alpine meadows, pastures, forest edges, scree slopes, nearby resting areas. | Look for the best compromise between high-quality forage, rapid escape opportunities and low disturbance. |
| Autumn | Territorial connectivity | Temporary reorganisation linked to the rut; sometimes long-distance movements. | Rutting grounds, convergence valleys, mountain passes, movement corridors. | Identify gathering areas and the corridors linking different sectors. |
| Winter | Zugänglichkeit | Reduzierte Bewegungen; Konzentration in energetisch günstigen Bereichen. | Talböden, südexponierte Hänge, schneearme Bereiche, untere Waldränder. | Auf zugängliche, geschützte und leicht schneebedeckte Bereiche nahe Deckung achten. |
| Frühling | Vernetzung + Ressourcen | Hohe Mobilität; Folgen von Schneeschmelze und Green-up; höhenbezogene und regionale Bewegungen. | Südhänge, Korridore, Pässe, Täler, Wald-Weide-Übergänge. | Erste schneefreie Bereiche identifizieren, die mit Rückzugsräumen verbunden sind. |
| Sommer | Ressourcen + Sicherheit | Relative Stabilisierung; lokale Nutzung von Äsungsflächen. | Alpine Matten, Weiden, Waldränder, Schutthalden, nahe Einstände. | Das beste Gleichgewicht zwischen hochwertigem Futter, raschem Rückzug und geringer Störung suchen. |
| Herbst | Territoriale Vernetzung | Vorübergehende Umorganisation durch die Brunft; teils weiträumige Bewegungen. | Brunftplätze, Konvergenztäler, Pässe, Bewegungskorridore. | Sammelräume und Korridore zwischen mehreren Sektoren erkennen. |
| Inverno | Accessibilità | Riduzione degli spostamenti; concentrazione nei settori meno costosi energeticamente. | Fondovalle, versanti esposti a sud, zone poco innevate, margini forestali bassi. | Cercare aree accessibili, riparate, poco innevate e vicine alla copertura. |
| Primavera | Connettività + risorse | Forte mobilità; inseguimento del disgelo e del green-up; movimenti altitudinali e regionali. | Versanti sud, corridoi, passi, valli, interfacce bosco-pascolo. | Individuare le prime zone libere dalla neve collegate a rifugi. |
| Estate | Risorse + sicurezza | Stabilizzazione relativa; uso locale delle aree di alimentazione. | Praterie alpine, pascoli, margini forestali, ghiaioni, aree di riposo vicine. | Cercare il miglior compromesso tra foraggio di qualità, possibilità di fuga rapida e basso disturbo. |
| Autunno | Connettività territoriale | Riorganizzazione temporanea legata al bramito; talvolta spostamenti a lunga distanza. | Aree di bramito, valli di convergenza, passi, corridoi di movimento. | Individuare le zone di raduno e i corridoi che collegano più settori. |
4. Différences spatiales entre sexes
Les mâles et les femelles n'utilisent pas l'espace de la même manière
Les observations de terrain suggèrent également que tous les individus n'utilisent pas le territoire de la même manière. Cette impression fait partie des tendances les plus constantes rencontrées dans les régions de montagne étudiées.
Dans la Combe de l'A, les femelles accompagnées de faons occupent principalement les altitudes intermédiaires, entre les forêts subalpines et les pâturages d'altitude. Les grands mâles sont plus souvent observés dans les pelouses alpines supérieures, les pierriers et les milieux minéraux proches des crêtes. Cette distribution n'est pas absolue, mais elle apparaît suffisamment souvent pour constituer une tendance récurrente.
Des observations similaires ont été réalisées dans le Val di Blenio. Sur plusieurs pâturages utilisés simultanément par les deux sexes, les groupes de femelles apparaissaient généralement plus tôt en soirée et restaient plus longtemps dans les milieux ouverts. Les mâles arrivaient souvent plus tard et utilisaient davantage les parties hautes ou périphériques des mêmes secteurs.
Au Marchairuz, les groupes de femelles se caractérisent aussi par une plus grande mobilité. Ils traversent régulièrement de vastes pâturages ouverts tout en maintenant un niveau élevé de vigilance collective. Les groupes observés présentent souvent une forte cohésion sociale et semblent s'appuyer sur une surveillance partagée. À plusieurs reprises, une femelle dominante occupait une position légèrement surélevée ou périphérique, jouant apparemment un rôle de sentinelle pour le reste du groupe.
Les mâles adultes suivent généralement une stratégie différente. Souvent observés seuls ou en groupes lâches, ils tendent à rester plus proches des zones de repos et apparaissent plus fortement associés aux structures de refuge du paysage. Dans plusieurs secteurs, les mâles ne s'aventuraient que partiellement dans les pâturages ouverts avant de regagner rapidement le couvert forestier ou l'abri topographique.
Ces observations sont globalement cohérentes avec la littérature scientifique. Alves et al. (2013) montrent que la ségrégation sexuelle est une caractéristique fondamentale du cerf hors période de rut. Conradt et al. (2000) suggèrent que les mâles adultes réagissent plus fortement aux conditions météorologiques défavorables, notamment au froid et au vent, et utilisent davantage les habitats abrités. Les femelles sont, quant à elles, davantage contraintes par les exigences énergétiques liées à la reproduction et à l'élevage des jeunes. Cette interprétation est renforcée par les travaux de Clutton-Brock et al. (1982), qui ont montré que les femelles allaitantes consacrent davantage de temps à l'alimentation que les femelles sans faon.
Il convient néanmoins de rester prudent. Les différences observées entre mâles et femelles ne doivent pas être interprétées comme des règles rigides. Des études récentes menées dans le Parc national suisse indiquent que les réponses aux ressources, au dérangement et à la structure du paysage peuvent varier selon les conditions locales. Comme souvent en écologie spatiale, les grandes tendances coexistent avec une variabilité individuelle et territoriale importante.
4. Spatial differences between sexes
Males and females do not use space in the same way
Field observations also suggest that not all individuals use territory in the same manner. This impression is among the most consistent patterns encountered across the mountain regions studied.
In the Combe de l'A, females accompanied by calves primarily occupy intermediate elevations between subalpine forests and high-altitude pastures. Large males are more frequently observed in upper alpine grasslands, scree slopes and mineral habitats located near ridgelines. This distribution is by no means absolute, but it appears sufficiently often to represent a recurring tendency.
Similar observations were made in the Val di Blenio. On several pastures used simultaneously by both sexes, groups of females generally appeared earlier in the evening and remained longer in open habitats. Males often arrived later and made greater use of higher or more peripheral sections of the same areas.
At Marchairuz, female groups are also characterised by greater mobility. They regularly traverse extensive open pastures while maintaining a high level of collective vigilance. Observed groups often display strong social cohesion and appear to rely on shared surveillance. On several occasions, a dominant female could be seen occupying a slightly elevated or peripheral position, apparently acting as a sentinel for the rest of the group.
Adult males generally follow a different strategy. Frequently observed alone or in loose groups, they tend to remain closer to resting areas and appear more strongly associated with refuge structures within the landscape. In several sectors, males ventured only partially into open pastures before rapidly returning to forest cover or topographic shelter.
These observations are broadly consistent with findings from the scientific literature. Alves et al. (2013) showed that sexual segregation is a fundamental characteristic of red deer outside the rutting season. Adult males and reproductive females often exploit different habitats and respond to distinct ecological constraints. Conradt et al. (2000) further suggested that adult males react more strongly to adverse weather conditions, particularly cold temperatures and wind, and therefore make greater use of sheltered habitats. By contrast, females are more constrained by the energetic demands associated with reproduction and calf rearing. This interpretation is reinforced by the classic work of Clutton-Brock et al. (1982), which demonstrated that lactating females devote more time to feeding than females without calves. Such differences in energetic requirements are likely to influence space use directly and may explain part of the contrasts observed in the field.
Nevertheless, caution remains necessary. Differences observed between males and females should not be interpreted as rigid rules. Recent studies conducted in the Swiss National Park indicate that responses to resources, disturbance and landscape structure may vary according to local conditions. As is often the case in spatial ecology, broad tendencies coexist with considerable individual and territorial variability.
4. Räumliche Unterschiede zwischen den Geschlechtern
Männchen und Weibchen nutzen den Raum nicht gleich
Feldbeobachtungen zeigen außerdem, dass nicht alle Individuen ein Gebiet auf dieselbe Weise nutzen. Dieser Eindruck gehört zu den konstantesten Mustern der untersuchten Gebirgsräume.
In der Combe de l'A halten sich Weibchen mit Kälbern vor allem in mittleren Höhenlagen zwischen subalpinen Wäldern und Hochweiden auf. Starke männliche Hirsche erscheinen häufiger in oberen alpinen Rasen, Schutthalden und mineralischen Bereichen nahe der Grate. Diese Verteilung ist nicht absolut, tritt aber häufig genug auf, um als wiederkehrende Tendenz betrachtet zu werden.
Ähnliche Beobachtungen wurden im Val di Blenio gemacht. Auf Weiden, die von beiden Geschlechtern genutzt wurden, erschienen Weibchengruppen meist früher am Abend und blieben länger in offenen Lebensräumen. Männchen trafen oft später ein und nutzten stärker die oberen oder randlichen Bereiche derselben Flächen.
Am Marchairuz zeichnen sich weibliche Rudel durch größere Mobilität aus. Sie durchqueren regelmäßig weite offene Weiden und bewahren dabei ein hohes Maß an kollektiver Wachsamkeit. Mehrfach konnte ein dominantes Weibchen in leicht erhöhter oder randlicher Position beobachtet werden, offenbar als Wächterin für die Gruppe.
Erwachsene Männchen verfolgen meist eine andere Strategie. Sie werden oft allein oder in lockeren Gruppen beobachtet, bleiben näher an Einständen und sind stärker an Rückzugsstrukturen der Landschaft gebunden. In mehreren Sektoren betraten Männchen offene Weiden nur teilweise, bevor sie rasch in Walddeckung oder topografischen Schutz zurückkehrten.
Diese Beobachtungen passen zu Alves et al. (2013) zur Geschlechtersegregation, zu Conradt et al. (2000) über Reaktionen auf ungünstige Wetterbedingungen und zu Clutton-Brock et al. (1982) über die erhöhten Nahrungsansprüche laktierender Weibchen.
Vorsicht bleibt dennoch wichtig. Unterschiede zwischen Männchen und Weibchen dürfen nicht als starre Regeln verstanden werden. Untersuchungen aus dem Schweizerischen Nationalpark zeigen, dass Reaktionen auf Ressourcen, Störung und Landschaftsstruktur von lokalen Bedingungen abhängen können. Allgemeine Tendenzen existieren neben erheblicher individueller und regionaler Variabilität.
4. Differenze spaziali tra i sessi
Maschi e femmine non usano lo spazio nello stesso modo
Le osservazioni di campo mostrano anche che non tutti gli individui utilizzano il territorio nello stesso modo. Questa impressione è una delle tendenze più costanti riscontrate nelle regioni montane studiate.
Nella Combe de l'A, le femmine accompagnate dai piccoli occupano soprattutto le quote intermedie, tra foreste subalpine e pascoli d'altitudine. I grandi maschi compaiono più spesso nei pascoli alpini superiori, nei ghiaioni e negli ambienti minerali prossimi alle creste. Questa distribuzione non è assoluta, ma appare abbastanza spesso da rappresentare una tendenza ricorrente.
Osservazioni analoghe sono state realizzate nel Val di Blenio. Su pascoli frequentati contemporaneamente dai due sessi, i gruppi di femmine comparivano generalmente prima la sera e rimanevano più a lungo negli spazi aperti. I maschi arrivavano spesso più tardi e sfruttavano maggiormente le parti alte o periferiche degli stessi settori.
Al Marchairuz, i branchi di femmine si distinguono per maggiore mobilità. Attraversano regolarmente ampi pascoli aperti mantenendo un elevato livello di vigilanza collettiva. I gruppi osservati mostrano spesso forte coesione sociale e sembrano affidarsi a una sorveglianza condivisa. In più occasioni è stata osservata una femmina dominante in posizione leggermente elevata o periferica, apparentemente con un ruolo di sentinella.
I maschi adulti seguono generalmente una strategia diversa. Spesso osservati soli o in gruppi poco coesi, tendono a restare più vicini alle aree di riposo e appaiono più legati alle strutture rifugio del paesaggio. In diversi settori si avventurano solo parzialmente nei pascoli aperti prima di tornare rapidamente alla copertura forestale o a un riparo topografico.
Queste osservazioni concordano con Alves et al. (2013) sulla segregazione sessuale, con Conradt et al. (2000) sulle risposte alle condizioni meteorologiche avverse e con Clutton-Brock et al. (1982) sui bisogni alimentari delle femmine in lattazione.
Resta tuttavia necessaria prudenza. Le differenze osservate tra maschi e femmine non devono essere interpretate come regole rigide. Studi recenti nel Parco Nazionale Svizzero indicano che le risposte alle risorse, al disturbo e alla struttura del paesaggio possono variare secondo le condizioni locali.


5. Lecture prédictive
Transformer une carte en hypothèse de présence
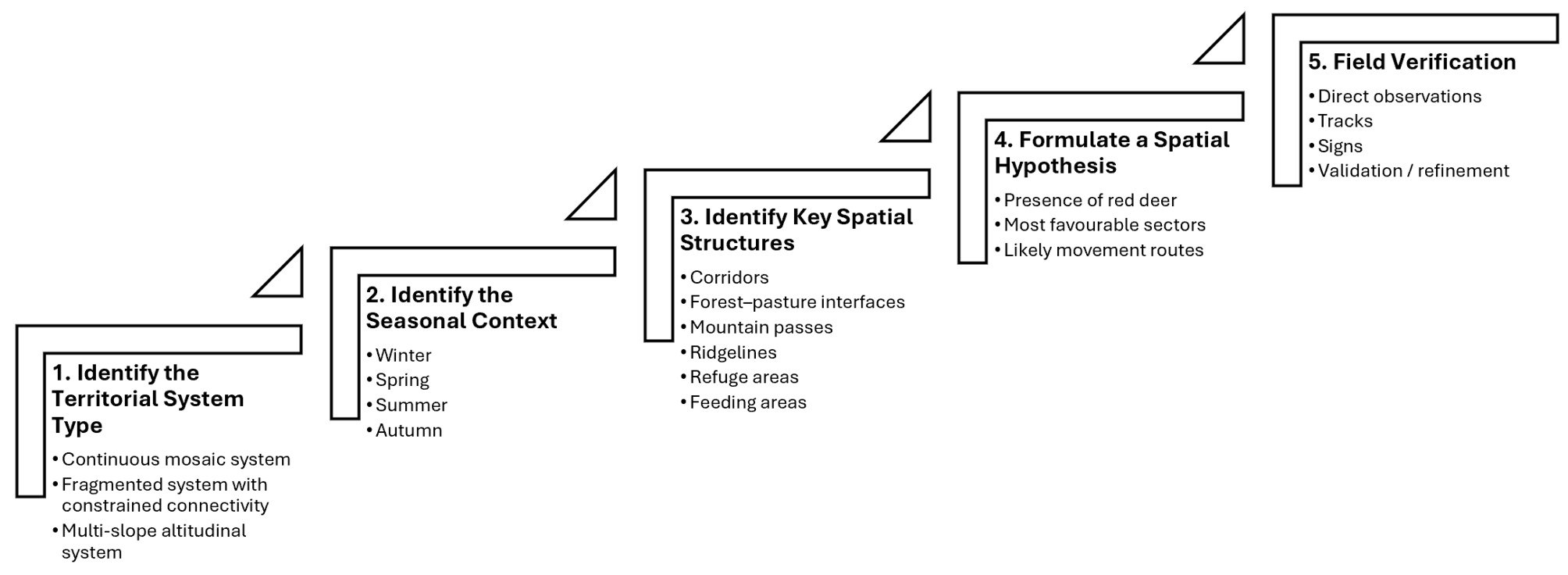
L'application pratique des modèles présentés ci-dessus repose sur une approche progressive de lecture du territoire. Cette approche consiste à passer d'une analyse générale du paysage à la formulation d'hypothèses de présence qui pourront ensuite être confrontées aux observations de terrain. La figure 4 résume les principales étapes de ce processus.
L'objectif n'est évidemment pas de prédire avec certitude la présence d'un animal sauvage. Il s'agit plutôt de hiérarchiser les différents secteurs d'un territoire selon leur probabilité d'utilisation et d'optimiser l'effort de prospection.
5. Predictive reading
From landscape interpretation to field prospecting: how to turn a map into a presence hypothesis
The practical application of the models presented above relies on a progressive approach to reading a territory. This approach consists of moving from a general analysis of the landscape to the formulation of presence hypotheses that can subsequently be confronted with field observations. Figure 4 summarises the main stages of this process.
The objective is obviously not to predict with certainty the presence of a wild animal. Rather, it is to rank the different sectors of a territory according to their probability of use and to optimise field prospecting efforts.
5. Vorausschauende Lesart
Aus einer Karte eine Präsenzhypothese entwickeln
Die praktische Anwendung der Modelle beruht auf einem schrittweisen Lesen des Gebiets. Von einer allgemeinen Landschaftsanalyse gelangt man zur Formulierung von Präsenzhypothesen, die anschließend im Gelände überprüft werden können. Abbildung 4 fasst die wichtigsten Schritte dieses Prozesses zusammen.
Ziel ist nicht, die Anwesenheit eines Wildtiers mit Sicherheit vorherzusagen. Vielmehr sollen die verschiedenen Bereiche eines Gebiets nach ihrer Nutzungswahrscheinlichkeit gewichtet und die Feldsuche effizienter gestaltet werden.
5. Lettura predittiva
Trasformare una carta in un'ipotesi di presenza
L'applicazione pratica dei modelli presentati si basa su un approccio progressivo alla lettura del territorio. Si passa da un'analisi generale del paesaggio alla formulazione di ipotesi di presenza, che vengono poi confrontate con le osservazioni sul campo. La figura 4 riassume le principali tappe del processo.
L'obiettivo non è prevedere con certezza la presenza di un animale selvatico. Si tratta piuttosto di classificare i settori di un territorio in base alla loro probabilità di utilizzo e di ottimizzare gli sforzi di prospezione.
Conclusion
Avant de chercher le cerf, apprendre à lire le territoire
Les observations présentées dans cet article suggèrent que le cerf élaphe n'occupe pas un territoire fixe, mais plutôt un système spatial dynamique dont l'organisation change constamment sous l'influence des saisons, de la disponibilité des ressources, des conditions météorologiques et de la structure sociale des groupes. La comparaison entre observations de terrain et littérature scientifique révèle plusieurs caractéristiques spatiales remarquablement constantes : interfaces entre milieux ouverts et couverts, corridors de déplacement, cols, gradients altitudinaux, zones refuges et noyaux d'habitat. Ces éléments apparaissent de manière répétée comme des facteurs clés structurant la distribution et les déplacements du cerf dans des paysages très variés.
La valeur principale d'une lecture géographique du paysage ne réside donc pas dans la recherche directe des animaux, mais dans la compréhension des structures qui rendent leur présence plus probable. Connectivité, accessibilité, distribution des ressources et sécurité forment ensemble un cadre spatial qui peut être interprété et utilisé pour guider la prospection de terrain.
Au-delà du cas du cerf élaphe, l'approche développée ici montre comment l'analyse du paysage peut contribuer à une compréhension plus large de la distribution de la faune. Les animaux deviennent non seulement des objets d'observation, mais aussi des indicateurs de l'organisation spatiale et du fonctionnement écologique des territoires qu'ils habitent.
Avant de chercher le cerf, il faut d'abord apprendre à lire le territoire.
Conclusion
Before looking for the deer, learn to read the territory
The observations presented in this article suggest that red deer do not occupy a fixed territory but rather a dynamic spatial system whose organisation constantly changes under the influence of seasons, resource availability, weather conditions and the social structure of groups. The comparison between field observations and scientific literature reveals a number of remarkably consistent spatial features: interfaces between open and covered habitats, movement corridors, mountain passes, altitudinal gradients, refuge areas and habitat cores. These elements repeatedly emerge as key factors shaping the distribution and movements of red deer across a wide variety of landscapes.
The principal value of a geographical reading of the landscape therefore lies not in searching directly for animals, but in understanding the structures that make their presence more likely. Connectivity, accessibility, resource distribution and safety together form a spatial framework that can be interpreted and used to guide field prospecting.
Beyond the specific case of red deer, the approach developed here illustrates how landscape analysis can contribute to a broader understanding of wildlife distribution. Animals become not only objects of observation but also indicators of the spatial organisation and ecological functioning of the territories they inhabit.
Before looking for the deer, one must first learn to read the territory.
Schlussfolgerung
Bevor man den Hirsch sucht, das Gebiet lesen lernen
Die Beobachtungen in diesem Beitrag zeigen, dass der Rothirsch kein festes Gebiet bewohnt, sondern ein dynamisches räumliches System nutzt, dessen Organisation sich unter dem Einfluss von Jahreszeiten, Ressourcenverfügbarkeit, Wetterbedingungen und sozialer Struktur der Gruppen ständig verändert. Der Vergleich zwischen Feldbeobachtungen und wissenschaftlicher Literatur zeigt bemerkenswert konstante Strukturen: Übergänge zwischen offenem und gedecktem Raum, Bewegungskorridore, Pässe, Höhengradienten, Rückzugsräume und Habitatkerne.
Der wichtigste Nutzen einer geografischen Landschaftslesung liegt daher nicht darin, Tiere direkt zu suchen, sondern die Strukturen zu verstehen, die ihre Anwesenheit wahrscheinlicher machen. Vernetzung, Zugänglichkeit, Ressourcenverteilung und Sicherheit bilden gemeinsam einen räumlichen Rahmen, der interpretiert und für die Feldsuche genutzt werden kann.
Über den Rothirsch hinaus zeigt dieser Ansatz, wie Landschaftsanalyse zu einem breiteren Verständnis der Wildtierverteilung beitragen kann. Tiere werden nicht nur zu Beobachtungsobjekten, sondern auch zu Indikatoren der räumlichen Organisation und des ökologischen Funktionierens ihrer Lebensräume.
Bevor man den Hirsch sucht, muss man zuerst lernen, das Gebiet zu lesen.
Conclusione
Prima di cercare il cervo, imparare a leggere il territorio
Le osservazioni presentate in questo articolo suggeriscono che il cervo non occupa un territorio fisso, ma un sistema spaziale dinamico la cui organizzazione cambia continuamente sotto l'effetto delle stagioni, della disponibilità di risorse, delle condizioni meteorologiche e della struttura sociale dei gruppi. Il confronto tra osservazioni di campo e letteratura scientifica mostra alcune caratteristiche spaziali notevolmente costanti: interfacce tra ambienti aperti e coperti, corridoi di movimento, passi, gradienti altitudinali, aree rifugio e nuclei di habitat.
Il valore principale di una lettura geografica del paesaggio non consiste quindi nel cercare direttamente gli animali, ma nel comprendere le strutture che rendono più probabile la loro presenza. Connettività, accessibilità, distribuzione delle risorse e sicurezza formano insieme un quadro spaziale che può essere interpretato e utilizzato per guidare la prospezione sul campo.
Al di là del caso specifico del cervo, l'approccio sviluppato qui mostra come l'analisi del paesaggio possa contribuire a una comprensione più ampia della distribuzione della fauna. Gli animali diventano non soltanto oggetti di osservazione, ma anche indicatori dell'organizzazione spaziale e del funzionamento ecologico dei territori che abitano.
Prima di cercare il cervo, bisogna imparare a leggere il territorio.
Références / References
Sources scientifiques et contextuelles
Scientific and contextual sources
Wissenschaftliche und kontextuelle Quellen
Fonti scientifiche e contestuali
- Alves, J., Støen, O.-G., Bischof, R., et al. (2013). Sexual segregation in red deer: patterns and hypotheses. Mammal Review, 43(3), 235-249.
- Clutton-Brock, T. H., Guinness, F. E., & Albon, S. D. (1982). Red Deer: Behaviour and Ecology of Two Sexes. Edinburgh University Press, Edinburgh.
- Conradt, L., Clutton-Brock, T. H., & Guinness, F. E. (2000). Costs and benefits of dispersion in female red deer. Animal Behaviour, 59, 451-459.
- Coppes, J., Ehrlacher, J., Thiel, D., & Braunisch, V. (2017). Outdoor recreation causes effective habitat reduction in capercaillie and other mountain wildlife. Journal of Applied Ecology, 54, 1197-1207.
- Federal Office for the Environment (FOEN). (2024). Federal Inventory of Swiss Wildlife Reserves. Bern.
- Federal Statistical Office (FSO). (2024). Swiss Land Use Statistics. Neuchâtel.
- Rempfler, V., Schmid, B., Signer, J., & Zweifel-Schielly, B. (2024). Resource selection and forage-safety trade-offs of red deer in alpine summer habitats. Movement Ecology, 12, 18.
- Rempfler, V., Schmid, B., Signer, J., & Zweifel-Schielly, B. (2025). Daytime habitat selection of red deer in the Swiss Alps: effects of vegetation cover, topography and recreation. Ecology and Evolution, 15, e70012.
- Sigrist, P., Coppes, J., Braunisch, V., & Bollmann, K. (2022). Green-up dynamics, landscape structure and habitat selection by red deer in mountain environments. Landscape Ecology, 37, 2041-2057.
- Urbina, J., Dufour, A.-B., Patthey, P., et al. (2023). Functional connectivity of red deer populations in western Switzerland: combining GPS telemetry and circuit theory. Landscape Ecology, 38, 2471-2488.
- State of Fribourg. (2024). Red Deer in the Canton of Fribourg: Distribution, Population Size and Management. Forests and Nature Service, Fribourg.
- State of Valais. (2025). High Hunting Season Report 2025. Wildlife, Hunting and Fisheries Service, Sion.
- Parco del Piano di Magadino. (2021). The Fauna of the Piano di Magadino Nature Park. Locarno.
- Swisstopo. (2024). swissALTI3D - Digital Elevation Model. Federal Office of Topography, Wabern.
- Swisstopo. (2024). swissTLM3D - Swiss Topographic Landscape Model. Federal Office of Topography, Wabern.
- Zweifel-Schielly, B., Kreuzer, M., Ewald, K. C., & Suter, W. (2009). Habitat selection by red deer in a topographically challenging alpine environment. European Journal of Wildlife Research, 55, 47-58.